


214-215
Организация клетки. Митохондрии
Транспортные системы
 Митохондрии имеют внутреннюю и внешнюю
мембраны (см. рис. 213). Внутренняя мембрана непроницаема для большинства
низкомолекулярных соединений. Она удерживает не только продукты промежуточного
метаболизма (например, пируват и ацетил-КоА), но и неорганические ионы
(Н+ и Na+). Поэтому в цитоплазме и митохондриях существуют
независимые пулы ионов и метаболитов. Напротив, внешняя мембрана содержит
порообразующие белки, которые делают ее проницаемой для низкомолекулярных
соединений (см. с. 212).
Митохондрии имеют внутреннюю и внешнюю
мембраны (см. рис. 213). Внутренняя мембрана непроницаема для большинства
низкомолекулярных соединений. Она удерживает не только продукты промежуточного
метаболизма (например, пируват и ацетил-КоА), но и неорганические ионы
(Н+ и Na+). Поэтому в цитоплазме и митохондриях существуют
независимые пулы ионов и метаболитов. Напротив, внешняя мембрана содержит
порообразующие белки, которые делают ее проницаемой для низкомолекулярных
соединений (см. с. 212).
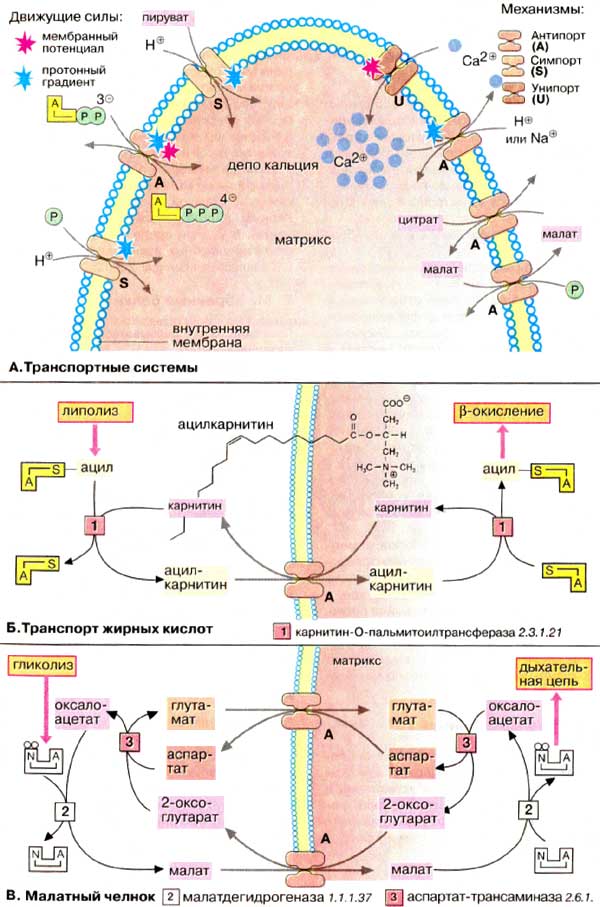
А. Транспортные
системы
Обмен между цитоплазмой и матриксом
обеспечивается специальными транспортными системами, локализованными во
внутренней мембране митохондрий и способными переносить разнообразные вещества
(пируват, фосфат, АТФ, АДФ, глутамат, аспартат, малат, 2-оксоглутарат, цитрат,
жирные кислоты) по механизмам типа антипорт (обменная диффузия,
А), симпорт (сопряженный транспорт, S) или унипорт
(облегченная диффузия, U) (см. рис. 221). Имеется переносчик и для ионов
Са2+, который наряду с ЭР регулирует концентрацию Са2+ в
цитоплазме.
Большая часть АТФ. продуцируемого
митохондриями в матриксе, доставляется в цитоплазму с помощью
АДФ/АТФ-транслоказы в обмен на АДФ (обменная диффузия). Фосфат поступает
в митохондрии вместе с протонами независимо от транспорта
АДФ/АТФ.
Аналогичным образом при участии
пируватспецифичного переносчика осуществляется одновременный перенос через
внутреннюю мембрану пирувата и протонов.
Б. Транспорт жирных
кислот
В митохондриях за перенос жирных кислот
отвечает специальная транспортная система. Активированные жирные кислоты в форме
ацил-КоА становятся транспортабельными в цитоплазме после взаимодействия с
карнитином. Образовавшийся ацилкарнитин транспортируется в матриксе
карнитиновым переносчиком, обмениваясь на свободный карнитин. В матриксе
ацильные остатки вновь связываются с КоА.
В. Малатный челнок
Для импорта восстановительных
эквивалентов в форме НАДН+Н+ (кофермент-связанного водорода),
образующихся в цитоплазме путем гликолиза, в митохондриях имеются несколько
челночных систем. В митохондриях млекопитающих этот транспорт осуществляется в
основном при помощи челночного механизма, использующего пару
малат-оксалоацетат. Основной функцией этого механизма является перенос
восстановительных эквивалентов в составе малата. Малат, попадая в матрикс
при посредстве переносчика, окисляется до оксалоацетата под действием
малатдегидрогеназы. Оксалоацетат переносится обратно в цитоплазму лишь после
трансаминирования в аспартат. Поскольку оксалоацетат может образовываться в
избыточном количестве, в реакции трансаминирования и последующем транспорте
принимает участие глутамат и 2-оксоглутарат. На схеме показано, что малатный
челнок функционирует в обоих направлениях, обеспечивая перенос восстановительных
эквивалентов от цитоплазматического НАДН в митохондрии без переноса
НАД+. В митохондриях насекомых трансмембранный перенос
восстановительных эквивалентов осуществляется с помощью глицерофосфатного
челнока.
Движущей силой транспортных процессов во
внутренней мембране митохондрий служит концентрационный градиент
метаболитов или электрохимический потенциал (см. рис. 143). Например,
карнитиновая система транспорта жирных кислот работает за счет высоких
концентраций ацил-КоА в цитоплазме. Движущей силой импорта фосфата и пирувата
служит протонный градиент, в то время как обмен АТФ/АДФ и выброс ионов
Са2+ зависят от трансмембранного потенциала внутренней
мембраны митохондрий.
Дополнительная
информация
Митохондрии являются главными
потребителями кислорода в организме. Кислородная недостаточность (гипоксия) как
результат недостаточного снабжения крови кислородом (ишемия) является причиной
повреждения тканей вплоть до некроза. Первым признаком гипоксии является
набухание митохондрий.