


368-369
Гормоны. Гидрофильные гормоны
Гидрофильные гормоны
 Гидрофильные гормоны и гормоноподобные
вещества построены из аминокислот. как, например, белки и пептиды, или являются
производными аминокислот. Они депонируются в больших количествах в клетках желез
внутренней секреции и поступают в кровь по мере необходимости. Большинство этих
веществ переносятся в кровотоке без участия переносчиков. Гидрофильные гормоны
действуют на клетки-мишени за счет связывания с рецептором на плазматической
мембране (см. с. 372).
Гидрофильные гормоны и гормоноподобные
вещества построены из аминокислот. как, например, белки и пептиды, или являются
производными аминокислот. Они депонируются в больших количествах в клетках желез
внутренней секреции и поступают в кровь по мере необходимости. Большинство этих
веществ переносятся в кровотоке без участия переносчиков. Гидрофильные гормоны
действуют на клетки-мишени за счет связывания с рецептором на плазматической
мембране (см. с. 372).
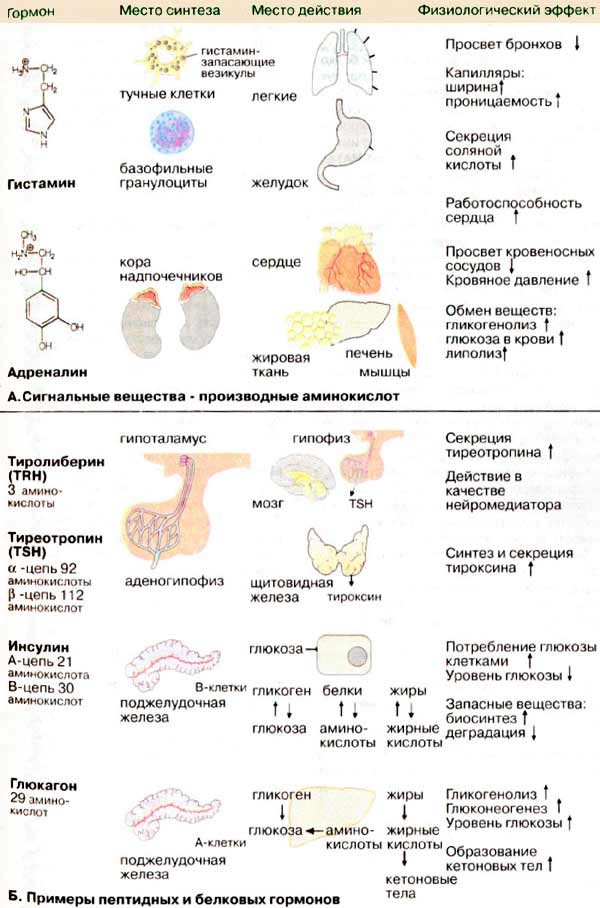
А. Сигнальные вещества — производные
аминокислот
Биогенные амины (гистамин,
серотонин, мелатонин) и катехоламины (дофа, дофамин, норадреналин и адреналин)
образуются путем декарбоксилирования аминокислот (см. с. 180).
Гистамин, важнейший медиатор
(локальное сигнальное вещество) и нейромедиатор, депонируется
главным образом в тучных клетках соединительной ткани и в базофильных
гранулоцитах крови. Он участвует в развитии воспалительных и аллергических
реакций. Освобождение гистамина происходит под действием веществ-либераторов,
таких, как тканевые гормоны, аллергены и лекарственные препараты. Действие
гистамина опосредовано различными типами рецепторов. Через H1-рецептор гистамин
стимулирует сокращение гладких мышц бронхов, расширяет капилляры и повышает их
проницаемость. Через Н2-рецептор гистамин замедляет сердечный ритм и стимулирует
образование соляной кислоты в желудочно-кишечном тракте. В центральной нервной
системе гистамин действует как нейромедиатор.
Адреналин — гормон коры
надпочечников, где он образуется из тирозина (см. с. 342). Выброс адреналина
находится под контролем центральной нервной системы. Как "аварийный гормон" он
воздействует главным образом на кровеносные сосуды, сердце и основной обмен.
Адреналин сужает сосуды и тем самым повышает кровяное давление (через
α1- и α2-рецепторы), повышает сердечную функцию (через
β1-рецепторы), ускоряет расщепление гликогена до глюкозы в печени и
мышцах и расширяет бронхи (через (β2-рецепторы).
Б. Примеры пептидных и белковых
гормонов
Эта самая большая группа сигнальных
веществ образуется в организме по обычному механизму белкового синтеза (см. с.
370). Низкомолекулярный пептидный гормон тиролиберин является трипептидом (362
Да). Высокомолекулярные белковые гормоны могут иметь молекулярную массу более 20
кДа, как, например, тиреотропин (28 кДа). Сходство первичной структуры некоторых
пептидных и белковых гормонов свидетельствует о том, что они относятся к
одному семейству и могли образоваться из одного эволюционного
предшественника.
Тиролиберин
[тиреотропин-рилизинг-фактор, ТРФ (TRH)], нейрогормон гипоталамуса (см. с. 342),
стимулирует секрецию клетками гипофиза тиреотропного гормона. ТРФ построен из
трех аминокислот, две из которых модифицированы (структура на с. 342).
N-концевая глутаминовая кислота присутствует в виде циклическою амида
(пироглутаминовой кислоты), а C-концевой пролин — в виде амида. Подобная
модификация делает молекулу устойчивой к действию экзопептидаз.
Тиреотропин [тиреотропный гормон,
ТТГ (TSH)] и родственные гормоны лютропин (лютеинизирующий гормон, ЛГ) и
фоллитропин (фолликулостимулирующий гормон, ФСГ) являются
представителями гормонов передней доли гипофиза. Они построены из двух
субъединиц и включают олигосахарид (являются гликопротеинами), который необходим
для быстрого удаления гормона из системы циркуляции. Тиреотропин стимулирует
синтез и выделение тироксина клетками щитовидной железы.
Инсулин (строение см. рис. 79) образуется
В-клетками поджелудочной железы и секретируется при повышении уровня глюкозы.
Роль инсулина в обмене веществ обсуждается на с. 162.
Глюкагон, пептидный гормон,
состоит из 29 аминокислотных остатков, синтезируется А-клетками (α-клетками
островков Лангерганса) поджелудочной железы. Глюкагон секретируется в кровь при
пониженном уровне глюкозы. Его основная функция состоит в повышении уровня
глюкозы (гипергликемический эффект) прежде всего за счет расщепления гликогена в
печени. По своему действию глюкагон является антагонистом
инсулина.