


154-155
Метаболизм углеводов
Гексозомонофосфатный путь
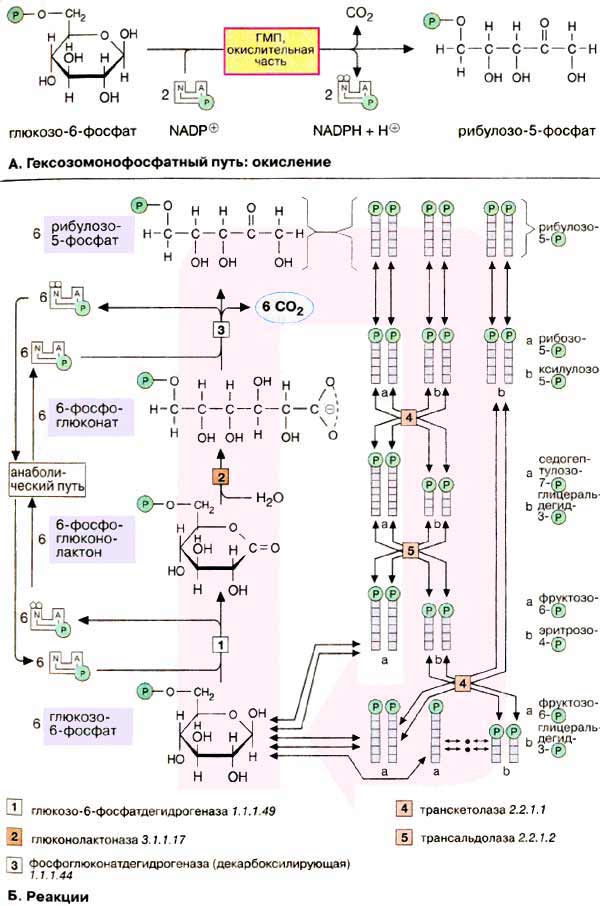 Гексозомонофосфатный путь [ГМП (HMW), часто называемый также
пентозофосфатным путем] является окислительным обменом веществ в цитоплазме,
в котором, как и в гликолизе, исходным субстратом служит глюкозо-6-фосфат.
ГМП поставляет два важных исходных соединения для анаболических процессов: НАДФН
+ Н+ (NADPH + Н+), необходимый для биосинтеза жирных кислот
и изопреноидов (см. с. 170), и рибозо-5-фосфат,
предшественник в биосинтезе нуклеотидов (см. с. 190).
Гексозомонофосфатный путь [ГМП (HMW), часто называемый также
пентозофосфатным путем] является окислительным обменом веществ в цитоплазме,
в котором, как и в гликолизе, исходным субстратом служит глюкозо-6-фосфат.
ГМП поставляет два важных исходных соединения для анаболических процессов: НАДФН
+ Н+ (NADPH + Н+), необходимый для биосинтеза жирных кислот
и изопреноидов (см. с. 170), и рибозо-5-фосфат,
предшественник в биосинтезе нуклеотидов (см. с. 190).
А. Гексозомонофосфатный путь:
окисление
В процессе окисления глюкозо-6-фосфат
превращается в рибулозо-5-фосфат. При этом образуются 1 молекула CO2
и 2 НАДФН + Н+. Значительно более сложная часть пути —
восстановительная (Б) — в зависимости от обмена веществ либо превращает часть
образованного пентозофосфата снова в гексозофосфат, либо включает его в гликолиз
для деградации. В большинстве клеток за счет ГМП разрушается не более 10%
глюкозо-6-фосфата.
Б. Реакции
Окислительная часть ГМП начинается с
окисления глюкозо-6-фосфата глюкозо-6-фосфатдегидрогеназой [1].
При этом образуется НАДФН + Н+ и 6-фосфоглюколактон —
внутримолекулярный сложный эфир (лактон) 6-фосфоглюконата. Специфическая
гидролаза (фермент [2]) расщепляет сложноэфирную связь и оставляет свободной
карбоксильную группу 6-фосфоглюконата. Последний фермент окислительной
части, фосфоглюконатдегидрогеназа (3), отщепляет карбоксильную группу
6-фосфоглюконата в виде CO2 с одновременным окислением гидроксильной
группы при С-3 до кетогруппы. Наряду со второй молекулой НАДФН + Н+
при этом образуется кетопентоза, рибулозо-5-фосфат, которая под действием
изомеразы превращается в рибозо-5-фосфат, исходное соединение для нуклеотидного
синтеза (на схеме сверху).
Восстановительная часть ГМП показана здесь только схематически.
Полная схема реакции представлена на с. 396.
Функция восстановительной ветви состоит
в том, чтобы производство НАДФН + Н+ и пентозофосфатов
соответствовало метаболическим потребностям клеток. Обычно потребность в НАДФН +
Н+ намного выше, чем в пентозофосфатах. В этих условиях 6 молекул
рибулозо-5-фосфата под действием трансальдолаз и транскетолаз образуют 5 молекул
фруктозо-6-фосфата, которые изомеризуются в 5 молекул глюкозо-6-фосфата.
Глюкозо-6-фосфат вновь участвует в окислительной части ГМП в процессе получения
НАДФН + Н+. Неоднократное повторение этих реакций позволяет окислить
глюкозо-6-фосфат до 6 молекул CO2. При этом образуется 12 молекул
НАДФН + Н+, а пентозофосфат не образуется.
При взаимном превращении фосфатов
сахаров в восстановительной части ГМП особенно важны два фермента.
Трансальдолаза [5] переносит С3-звенья от
седогептулозо-7-фосфата, кетосахара с 7 атомами углерода, на альдегидную группу
глицеральдегид-3-фосфата. Аналогичным образом транскетолаза [4] катализирует
перенос С2-фрагмента с одного фосфата сахара на
другой.
Реакции восстановительной части ГМП
обратимы, т.е. гексозофосфаты могут непосредственно превращаться в
пентозофосфаты. Это превращение может происходить при высокой потребности клетки
в пентозофосфатах, например на стадии репликации ДНК и S-фазе клеточного цикла
(см. с. 380).
Дополнительная
информация
Если наряду с НАДФН + Н+
клетке требуется энергия в форме АТФ, продукты восстановительной части ГМП
(фруктозо-6-фосфат и глицеральдегид-3-фосфат) включаются в гликолиз и далее в
цитратный цикл и дыхательную цепь с образованием CO2 и воды. На этом
пути из 6 молей глюкозо-6-фосфата образуется 12 молей НАДФН + Н+ и
примерно 150 молей АТФ.