


156-157
Метаболизм углеводов
Глюконеогенез
 Некоторые ткани, такие, как мозг и эритроциты, зависят
от постоянного снабжения глюкозой. Если получаемое с пищей количество углеводов
недостаточно, необходимая концентрация глюкозы в крови может поддерживаться
некоторое время за счет расщепления гликогена печенью (см. с. 158).
Если истощены также и эти запасы, в печени запускается синтез глюкозы de
novo, глюконеогенез (см. с. 302). Наряду с
печенью высокой глюконеогенезной активностью обладают также клетки
почечных канальцев (см. с. 320). Исходными соединениями
в глюконеогенезе являются аминокислоты мышечной ткани. При длительном
голодании это приводит к массивному распаду мышечного белка. Другими важными
исходными веществами для синтеза глюкозы служат лактат, образующийся
в эритроцитах и мышечной ткани при недостатке О2 , а также глицерин,
образующийся при расщеплении жиров. Напротив, жирные кислоты не могут трансформироваться
в глюкозу в организме животных, так как в данном случае деградация жирных кислот
не является анаплеротическим процессом (см. с. 140). В
организме человека за счет глюконеогенеза образуется несколько сотен граммов
глюкозы в сутки.
Некоторые ткани, такие, как мозг и эритроциты, зависят
от постоянного снабжения глюкозой. Если получаемое с пищей количество углеводов
недостаточно, необходимая концентрация глюкозы в крови может поддерживаться
некоторое время за счет расщепления гликогена печенью (см. с. 158).
Если истощены также и эти запасы, в печени запускается синтез глюкозы de
novo, глюконеогенез (см. с. 302). Наряду с
печенью высокой глюконеогенезной активностью обладают также клетки
почечных канальцев (см. с. 320). Исходными соединениями
в глюконеогенезе являются аминокислоты мышечной ткани. При длительном
голодании это приводит к массивному распаду мышечного белка. Другими важными
исходными веществами для синтеза глюкозы служат лактат, образующийся
в эритроцитах и мышечной ткани при недостатке О2 , а также глицерин,
образующийся при расщеплении жиров. Напротив, жирные кислоты не могут трансформироваться
в глюкозу в организме животных, так как в данном случае деградация жирных кислот
не является анаплеротическим процессом (см. с. 140). В
организме человека за счет глюконеогенеза образуется несколько сотен граммов
глюкозы в сутки.
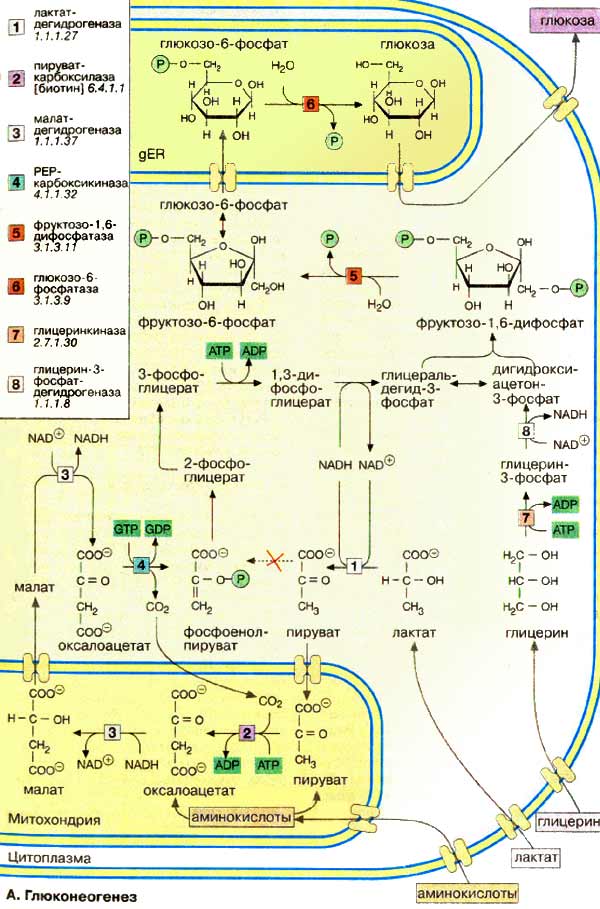
А. Глюконеогенез
Многие реакции глюконеогенеза
катализируются теми же ферментами, что и процессы гликолиза (см. с. 152).
Некоторые ферменты специфичны для глюконеогенеза и синтезируются только по мере
необходимости под воздействием кортизола и гпюкагона (см. с. 160).
На схеме представлена только эта группа ферментов. В то время как гликолиз
протекает в цитоплазме, глюконеогенез происходит также в митохондриях и
эндоплазматическом ретикулуме.
Первые стадии реакционной цепи протекают в митохондриях.
Причиной такого «обходного» пути является неблагоприятная константа равновесия
пируваткиназной реакции (см. с. 152). Для перевода пирувата
непосредственно в фосфоенолпируват(PEP) недостаточно энергии расщепления
АТФ. Пируват, образующийся из лактата или аминокислот, переносится в
матрикс митохондрий и там карбоксилируется в оксалоацетат в биотинзависимой
реакции, катализируемой пируваткарбоксилазой [2].
Оксалоацетат является промежуточным метаболитом цитратного цикла. Поэтому
аминокислоты, которые включаются в цитратный цикл или конвертируются в пируват,
могут непосредственно превращаться в глюкозу (глюкогенные аминокислоты, см.
с. 182).
Оксалоацетат, образующийся в митохондриальном матриксе, восстанавливается
в малат [3]. который может переноситься
в цитоплазму с помощью специальных переносчиков (см. с. 214).
Оксалоацетат может также переноситься из митохондрии в цитоплазму после переаминирования
в аспартат (малатный челночный механизм, см. с. 206).
В цитоплазме малат вновь превращается
цитоплазматической малатдегидрогеназой в оксалоацетат, который в реакции,
катализируемой ГТФ-зависимой РЕР-карбоксикиназой [4], переводится в
фосфоенолпируват. Последующие стадии до фруктозо-1,6-дифосфата
представляют собой модификации соответствующих реакций гликолиза. При этом для
образования 1,3-дифосфоглицерата дополнительно расходуется АТФ.
Две глюконеогенез-специфичные
фосфатазы отщепляют по очереди фосфатные остатки от
фруктозо-1,6-дифосфата. Промежуточной стадией является изомеризация
фруктозо-6-фосфата в глюкозо-6-фосфат, одна из реакций гликолиза.
Глюкозо-6-фосфатаза печени [5] является мембранным ферментом,
локализованным внутри гладкого эндоплазматического ретикулума. Перенос
глюкозо-6-фосфата в эндоплазматический ретикулум и возврат образующейся
глюкозы в цитоплазму осуществляется специфическими переносчиками. Из
цитоплазмы глюкоза поступает в кровь.
Глицерин прежде всего
фосфорилируется [7] в положении 3. Образующийся 3-глицерофосфат окисляется
НАД+-зависимой дегидрогеназой [8] в дигидроксиацетон-3-фосфат,
который далее включается в глюконеогенез.