


140-141
Метаболизм. Энергетика
Цитратный цикл: метаболические функции
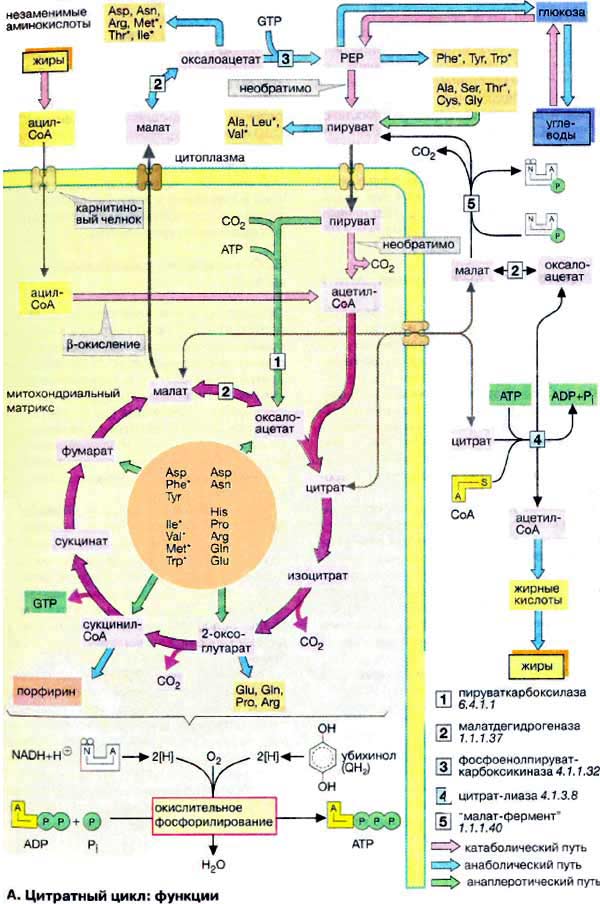 Цитратный цикл:
функции
Цитратный цикл:
функции
Цитратный цикл (см. с. 138) играет
центральную роль в промежуточном метаболизме клетки. Наряду с катаболическими и
анаболическими цикл выполняет и амфиболические функции. Промежуточные
соединения цитратного цикла, включая такие важные метаболиты, как пируват и
ацетил-КоА, способные окисляться до CO2, идентичны промежуточным
соединениям многих катаболических путей. Образующиеся в цикле восстановительные
эквиваленты (см. с. 20) окисляются в дыхательной цепи (окислительное
фосфорилирование) с образованием АТФ (АТР) (см. ниже).
Промежуточные соединения цитратного
цикла включаются во многие процессы биосинтеза, например в биосинтез
глюкозы (глюконеогенез; оксалоацетат и малат), синтез
порфиринов (сукцинил-КоА) и синтез аминокислот (2-оксоглутарат,
оксалоацетат). Кроме того, цитратный цикл поставляет в цитоплазму
ацетил-КоА, необходимый для синтеза жирных кислот и
изопреноидов.
Ацетил-КоА, образующийся в матриксе митохондрий при участии
пируватдегидрогеназы (см. с. 136), не может проходить
через внутреннюю митохондриальную мембрану. Поэтому ацетильный остаток конденсируется
митохондриальной цитрат-синтазой с оксалоацетатом с образованием цитрата.
Последний переносится в цитоплазму по механизму антипорта с малатом (см. с.
214), где снова расщепляется АТФ-зависимой цитрат-лиазой
[4] с образованием ацетил-КоА и оксалоацетата.
Образовавшийся оксалоацетат восстанавливается цитоплазматической малатдегидрогеназой
[2] в малат, который возвращается в митохондрии
за счет антипорта или подвергается окислительному декарбоксилированию «малат-ферментом»
[5] с образованием пирувата. Образующийся НАДФН
+ Н+ принимает участие в биосинтезе жирных кислот.
Промежуточные продукты цитратного цикла
присутствуют в митохондриях лишь в очень незначительных количествах. При
окислении ацетил-КоА они вновь регенерируются, так что их концентрации остаются
практически постоянными. В то же время анаболические процессы быстро истощают
пул некоторых промежуточных продуктов цикла. Поэтому их запас постоянно
пополняется за счет метаболитов, поступающих из других источников.
Ферментативные процессы, пополняющие запас промежуточных продуктов цикла,
называются анаплеротическими (возмещающими) реакциями (см. с.
168).
Анаплеротический характер носит деградация большинства аминокислот,
так как при этом образуются промежуточные соединения цикла или пируват (глюкогенные
аминокислоты, см. с. 156). Фактически глюконеогенез поддерживается
в основном за счет деградации аминокислот. Особенно важной анаплеротической
стадией в метаболизме животных является превращение пирувата в оксалоацетат.
Эта АТФ-зависимая реакция, катализируемая пируваткарбоксилазой [1],
позволяет включать в глюконеогенез пируватпоставляющие аминокислоты и лактат.
В отличие от пирувата ацетил-КоА не
является анаплеротическим метаболитом у высших животных. Его углеродный скелет
полностью окисляется до CO2 и поэтому не принимает участия в
биосинтезе. Поскольку при деградации жирных кислот образуется ацетил-КоА, клетки
животных не в состоянии превращать жирные кислоты в глюкозу. Поэтому при
голодании в организме прежде всего утилизируются не жиры, а белки.
Высвободившиеся аминокислоты, напротив, могут превращаться и в жирные кислоты, и
в глюкозу и, тем самым, поддерживать уровень сахара в крови (см. с.
300).
Дополнительная информация
В растениях и бактериях ацетил-КоА
превращается в сукцинат в так называемом глиоксилатном цикле, тесно
связанном с цитратным циклом. Эти организмы способны осуществлять
анаплеротическую деградацию нейтральных жиров. В растениях глиоксилатный путь
локализован в особых органеллах, глиоксисомах.